
Part1cGAS-STING驱动与衰老相关的炎症和神经变性


通讯作者

1背景
衰老的特点是机体适应性下降,并增加了对各种疾病的易感性。尽管多种稳态机制的功能受损可以独立地导致衰老过程,但其中许多机制会聚集产生异常的炎症状态,从而导致与年龄相关的衰退。事实上,与年龄有关的炎症的减弱已成为一种常见的机制,通过这种机制,(药物)干预对衰老发挥其有益作用。
炎症通常是由先天免疫系统的模式识别受体的参与引发的。之前已有研究描述了cGAS-STING途径在调节细胞衰老中的作用——衰老的标志(参考:https://mp.weixin.qq.com/s/fOhVoso7dNCagnmu4NvfIQ)。然而,cGAS-STING途径是否直接导致人体组织中的细胞衰老或体内与年龄相关的炎症和功能障碍尚不清楚。
2结果
STING引发与年龄相关的炎症
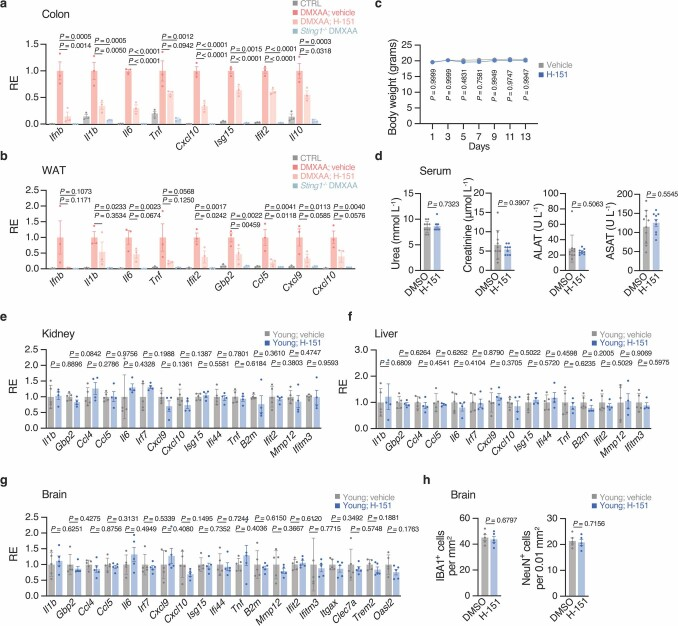
a, b与DMXAA处理或未处理(CTRL)的WT和Sting1−/−小鼠(对照),结合载药或H-151 (n = 3),相对于DMXAA载药处理小鼠的平均诱导水平,测量结肠(a)和白色脂肪组织(WAT, b)中促炎和干扰素相关基因的mRNA表达水平。结肠:H-151 P = 3×10^−6(白细胞介素6),8×10^−5 (Cxcl10), 9×10^−7 (Isg15), 8×10^−5 (Ifit2);Sting1−−/ P = 2×10^−7(白细胞介素6),4×10^−6 (Cxcl10), 5×10^−8 (Ifit2)。
c-h,幼龄小鼠每天用H-151治疗14天。
c,给药或H-151处理小鼠体重随时间变化(n = 10)。
d, H-151处理小鼠(n = 9)和未处理小鼠(n = 10)的血清肌酐、尿素、ALAT和ASAT水平。
e - g, H-151处理幼鼠(肾、肝n = 4,脑n = 5)和未处理幼鼠(n = 5)肾脏(e)、肝脏(f)和脑(g)中促炎和干扰素相关基因mRNA表达水平。相对表达量与DMXAA处理小鼠的平均诱导水平比较。
h,未注射H-151的幼鼠海马IBA1小胶质细胞(左)和NeuN神经元(右)的计数(n = 5)。
数据为平均值±s.e.m。P值的计算采用单侧普通方差分析,然后进行Tukey多重比较检验(a,b),采用双侧Student’s unpaired t检验(d-h)或采用Geisser-Greenhouse校正的RM双向方差分析,然后进行Sidak多重比较检验(c)。扩展图(Extended Data Fig)和Supplementary figure都用于展示额外的图表数据,
"Extended Data Fig"是一种特定于《自然》杂志(Nature)的图表形式,
这些图表通常通过在线版的文章进行展示,而不是出现在打印版中。DMXAA: mSTING激动剂; WAT: white adipose tissue(白色脂肪组织); vehicle组:是指使用一种无活性药物或溶剂来进行对照处理。
为了研究cGAS-STING通路在年龄相关表型中的作用,我们首先测试了选择性和耐受性良好的小分子抑制剂H-151对STING的急性抑制是否可以抑制衰老细胞的炎症反应,这是研究年龄相关炎症的一种范式体外模型(扩展图1)。延伸先前的研究, H-151介导的STING抑制在各种衰老情况下有效抑制了几种促炎基因和I型干扰素(IFN)刺激基因(ISGs)的诱导,而不影响衰老细胞的其他非炎症特征(图1a,b和扩展图2a-e)。
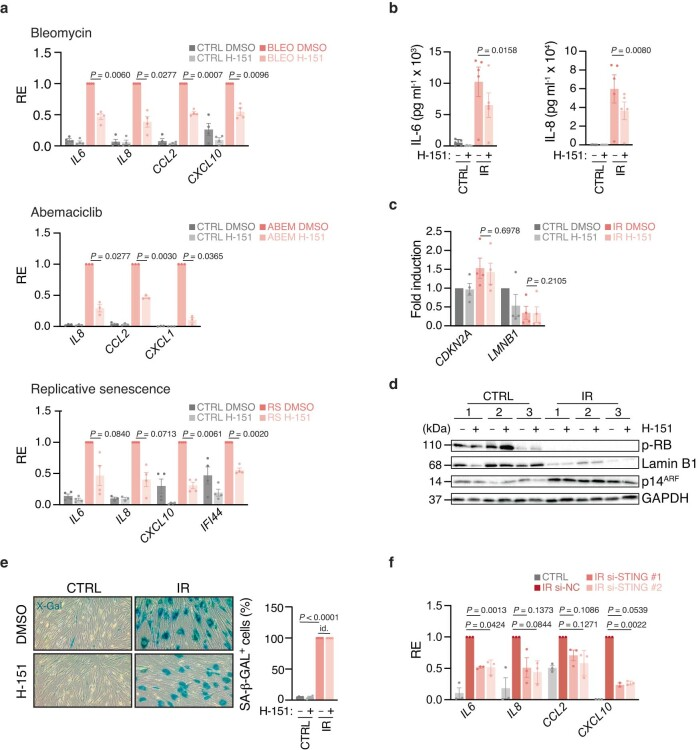
a,分别用DNA代谢抑制剂博莱霉素(n = 4个实验)、CDK4/6抑制剂Abemaciclib (n = 3个实验)、重复传代(n = 4个实验)或在5% O2 (CTRL)中维持,然后用DMSO或H-151(每天0.5 μM,持续10天)处理后衰老的人WI-38成纤维细胞的促炎基因和干扰素刺激基因的mRNA表达水平,在每个实验中相对于DMSO刺激条件下的诱导水平进行测量。
b,在DMSO或H-151处理的对照和辐照WI-38细胞培养液(每天0.5 μM,从第10天到第20天,共n = 5个实验)中,通过ELISA测定细胞因子产量。
c, b中处理的细胞中衰老特征基因的mRNA表达水平(n = 4个实验),在每个实验中相对于对照DMSO条件测量。
d,第20天收获的(b)中处理过的辐照细胞的Western blot表征(n = 3个实验)。
e,对照和照(b)处理的WI-38细胞衰老相关ß-半乳糖苷酶(SA-ß-GAL)染色的代表性图像和定量,相对于DAPI细胞总数测量(n = 4 FOV,代表n = 3个实验),P = 4 × 10^−5。Id:相同的值。
f, si-NC或si-STING处理的对照和辐照(12 Gy, IR)人BJ成纤维细胞(照射后72小时,10天)促炎和干扰素刺激基因的mRNA表达水平。每个实验(n = 3)的相对表达式归一化到辐照si-NC条件下的感应。
数据为平均值±s.e.m。P值采用双侧配对比Student’s t检验(a-c, e,f)。WI-38细胞是一种人类胚胎肺成纤维细胞系。CDKN2A是肿瘤抑制基因,参与细胞增殖调控;LMNB1是细胞核骨架蛋白质基因,参与细胞核结构和功能的维持。p-RB参与细胞周期调控;Lamin B1维持细胞核结构和功能;p14调控p53蛋白的稳定性和活性;GAPDH参与代谢途径并具有其他细胞功能。(扩展图2c-e 说明了H-151不影响衰老细胞的其他非炎症特征)
与H-151的药理抑制相比,在完全衰老细胞中靶向STING的RNA干扰产生了类似的结果(扩展图2f)。
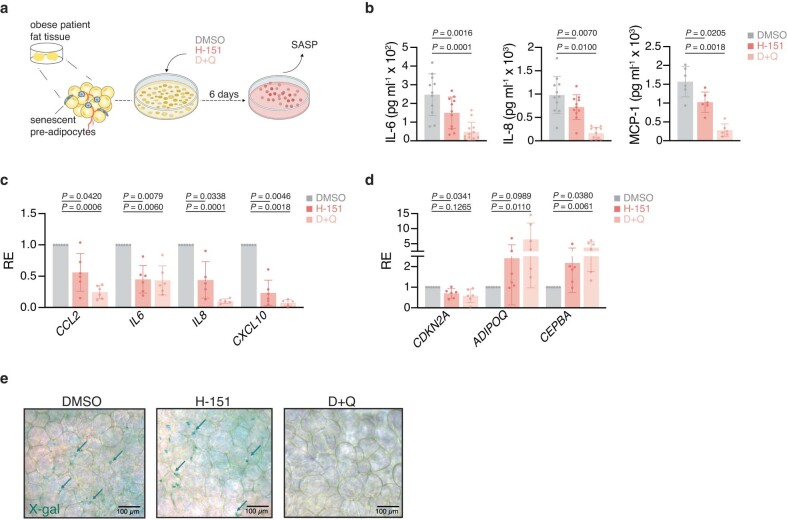
a,大网膜脂肪组织移植研究示意图。
b, ELISA法测定个体患者组织外植体条件培养基中IL-6 (n = 11)、IL-8 (n = 11)和MCP-1 (n = 6)的含量。
c,d,促炎和干扰素刺激基因(c)和脂肪组织功能基因(d)的mRNA表达水平,每个患者(n = 6)归一化到DMSO对照条件。
e,大网膜脂肪组织衰老相关β-半乳糖苷酶(SA-β-GAL)染色的代表性图像(n = 1个组织,代表n = 11例患者),用RapiClear清除。箭头表示SA-β-GAL细胞。标尺,100 μm。
数据为平均值±s.e.m。P值采用双侧配对比Student’s t检验(b-d)。D + Q:抗衰老药物达沙替尼与槲皮素联合用药MCP-1(单核细胞趋化蛋白-1)是一种细胞因子, 参与炎症反应和免疫调节过程。RapiClear是一种组织清理剂,可用于去除标本中的油脂、蜡和其他污染物,使组织标本更清晰、透明,并提供更好的显微镜观察和分析。D+Q:
https://mp.weixin.qq.com/s/KVJWUsw7s7sF4jZYIoplqg
使用肥胖个体的人体脂肪组织外植体,其中脂肪组织积累了衰老的前脂肪细胞,我们证实了H-151抑制STING在组织水平上抑制衰老细胞释放促炎信号(扩展图3)。
这些数据表明,抑制细胞和人体组织中的STING可以阻断衰老细胞的炎症反应,衰老细胞是衰老过程中炎症的主要原因。
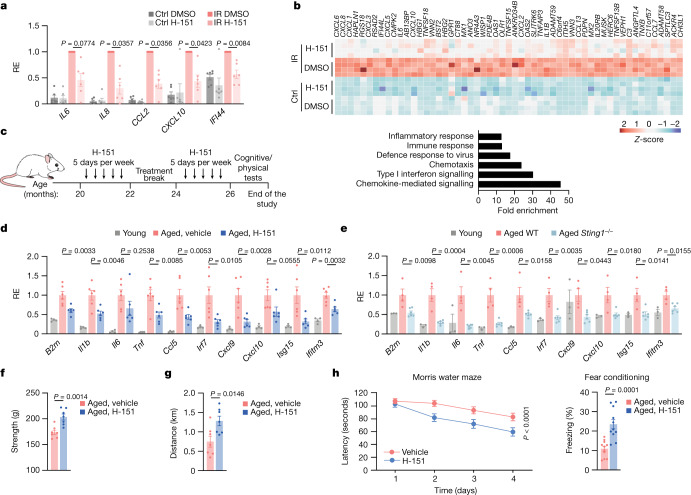
a,b,人WI-38成纤维细胞的促炎基因和ISGs mRNA表达水平(a)和RNA-seq分析(b),分别在辐照(12 Gy, IR)或维持在5% O2 (Ctrl)下,H-151(每天,0.5 μM)或DMSO处理10天后衰老(第10天至20天)。测量每个实验(n = 6)相对于辐照DMSO条件下诱导水平的相对表达量(RE) (a)。
b,辐照后表达最多、H-151处理后抑制最多的前50个基因(n = 4个实验)(上),以及基于上述基因列表的基因集富集分析(下)。
c,与d和f-h所示数据相关的H-151对野生型(WT)老龄小鼠的处理示意图。
d,e, H-151处理或不处理的年轻小鼠(n = 4)和老年小鼠(n = 6) (d)以及年轻小鼠(n = 3)、老年WT (n = 4)和Sting1 - / -小鼠(n = 6) (e)中促炎基因和ISGs的肾脏mRNA表达水平。相对于老龄小鼠(d)或老年WT (e)的平均值,测量了表达水平。
f,g,给予H151或不给予H-151的老年小鼠(n = 7)的身体状况,通过握力(f)和跑步机跑步距离(g)来评估。
h,认知功能测试(n = 11只小鼠)使用Morris水迷宫测试(左图,多日到达平台的延迟时间)和恐惧条件反射(右图,冻结时间百分比)来评估。P = 3 × 10^−5。
数据为平均值±s.e.m。P值采用双侧配对比例学生t检验(a)、双侧未配对学生t检验(f-h)(右)、单因素方差分析(ANOVA),然后进行Tukey多重比较检验(d和e)和普通双因素方差分析(h,左)。接下来,我们使用H-151阻断老龄小鼠的STING(图1c)。与先前的研究一致,在非常大的年龄(26个月),小鼠在肾脏和肝脏中显示出炎症和I型IFN特征(图1d,扩展图4a,b)。值得注意的是,由于STING抑制,各种与衰老相关的免疫特征基因显著减弱(图1d,扩展图4a,b)。同样,与老龄野生型小鼠相比,老龄Sting1−/−小鼠显示出衰老相关免疫标志物水平下降,独立验证了H-151在小鼠体内研究STING作用的有效性(图1e,扩展图4f)。接下来,我们研究了STING依赖性炎症的衰减是否会影响老年小鼠的身体和认知功能。与对照组小鼠相比,H-151处理的老年小鼠的肌肉力量和身体耐力(分别通过握力和跑步机测量)均有所改善(图1f,g)。在测试海马依赖学习和记忆表现时,我们观察到STING抑制导致Morris水迷宫测试中的空间记忆和情境恐惧条件反射测试中的联想记忆显著改善(图1h)。
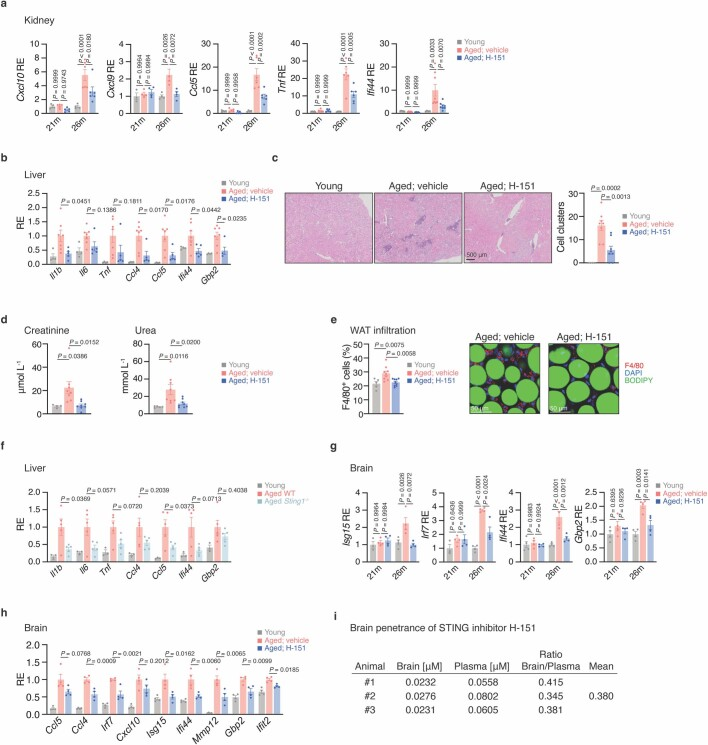
a,幼龄小鼠(21m n = 5 -Cxcl9 n = 3-, 26m n = 4)、21和26个月H-151处理小鼠(21m n = 5 -Cxcl9 n = 4-, 26m n = 6 -Cxcl9 n = 4, Ifi44 n = 8-)和未处理小鼠(21m n = 5 -Cxcl9 n = 4-, 26m n = 6 -Cxcl9 n = 4, Ifi44 n = 8-)肾脏促炎和干扰素刺激基因mRNA表达水平。P = 3 × 10^-5 (Cxcl10), 3×4×10^−7 (Ccl5), 2×10^−7 (Tnf)。
b,年轻小鼠(n = 4)和老龄小鼠(n = 5) H-151处理和未处理H-151处理小鼠(n = 7)肝脏促炎和干扰素刺激基因mRNA表达水平。
c,年轻小鼠(n = 4)和未处理H-151的老年小鼠(n = 8)肾脏炎症团的代表性组织学图像和量化。比例尺,500 μm。
d,年轻(n = 4)和老年(n = 8)小鼠H-151处理前后血清肌酐(左)和尿素(右)水平。
e,相对于DAPI细胞,H-151处理或未处理的年轻小鼠(n = 5)和老年小鼠(n = 9)白色脂肪组织(WAT)中F4/80巨噬细胞(红色)的代表性图像和定量。标尺,50 μm。
f,幼年(n = 3)和老年WT/Sting1++−/−小鼠(n = 5)肝脏促炎和干扰素刺激基因mRNA表达水平,归一化为老年小鼠平均值。
g,年轻小鼠(21m n = 4 - Isg15, Irf7 n = 3-, 26m n = 4)和未处理H-151的21和26个月小鼠(n = 4)脑促炎和干扰素刺激基因mRNA表达水平。P = 4 × 10^−6 (Irf7), 6 × 10^−5 (Ifi44)。
h, H-151处理或未处理的年轻/老年小鼠脑促炎和干扰素刺激基因mRNA表达水平(n = 4),归一化为老龄小鼠的平均水平。
i,小鼠H-151的脑外显率测量。
数据为平均值±s.e.m。P值采用单因素方差分析计算,然后采用Tukey多重比较检验(a-h)。在炎症细胞因子减少的同时,STING抑制的动物显示肾脏炎症细胞积聚减少,与药物处理的老年小鼠相比,肾脏损伤标志物(肌酐、尿素)水平降低,白色脂肪组织中巨噬细胞数量减少(扩展图4c-e)。
一致地,H-151(一种脑渗透化合物)抑制STING可降低老年小鼠大脑中免疫相关特征基因的水平(扩展图4g-i)。
总之,这些结果表明STING是衰老相关炎症的重要驱动因素,无论是在外周还是中枢神经系统,都会促进衰弱(frailty)和认知能力下降。
cGAS-STING影响大脑衰老
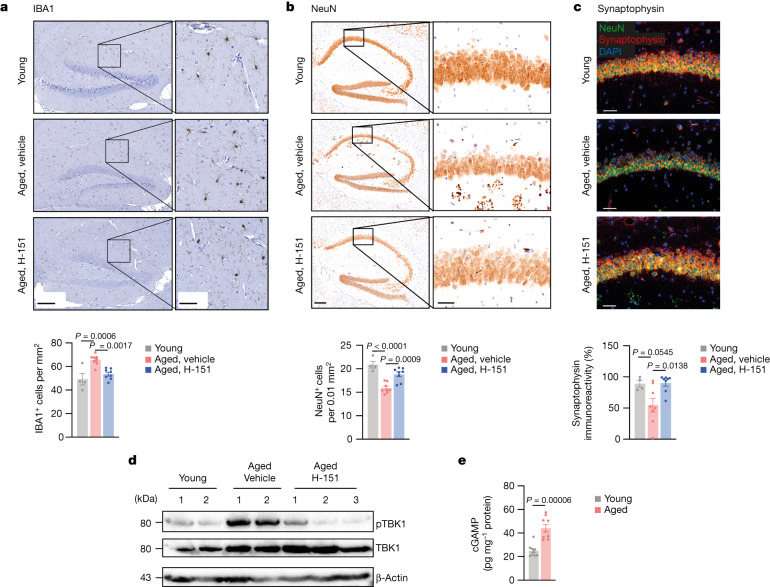
a - c,经H-151处理或未处理的幼龄小鼠(n = 4)和老年小鼠(n = 8)脑切片海马IBA1细胞(a)、NeuN细胞(b)和CA1区突触素强度(c)的代表性图像和定量。比例尺:200 μm (a、b(左))、50 μm (a、b(右)、c)。P = 3 × 10^−5。
d, Western blot分析幼龄小鼠(n = 2)、老年小鼠(n = 2)和H-151急性治疗老年小鼠(每天连续5天,n = 3)脑裂解物中pTBK1的含量。
e,用酶联免疫吸附法(ELISA)测定青年和老年小鼠脑裂解物中cGAMP的产生(n = 9)。
数据为平均值±s.e.m。P值的计算采用普通的单因素方差分析,然后采用Tukey多重比较检验(a-c)或双侧未配对学生t检验(e)。在老化的大脑中,引发不适应炎症和I型IFN反应的信号机制尚不完全清楚。因此,我们专注于了解cGAS-STING通路对大脑衰老的贡献。老年小鼠海马实质组织病理学检查显示小胶质细胞增生,STING抑制后小胶质细胞增生减少(图2a)。
此外,STING抑制可以保护小鼠免受海马CA1区域神经元的损失,并增加突触素(突触活性的标志物)的局部水平(图2b,c)。
在衰老过程中,系统性因素会损害成人神经发生并导致认知功能障碍。为了确定所观察到的效应是否由脑内STING活性引起,我们分析了老年小鼠全脑裂解物中STING信号的生物标志物。磷酸化TBK1 (Ser172;pTBK1) ——负责STING19,20下游信号转导的主要激酶——在老年小鼠大脑中表达增加(图2d)。老年小鼠急性STING抑制导致pTBK1信号减少,表明STING-TBK1信号轴局部参与老年脑(图2)。在DNA感知(DNA sensing)中,STING被cGAS内源性产生的2’3′-cGAMP(cGAMP)激活。我们在老年小鼠的脑裂解物中检测到强大的cGAMP产生,但在年轻小鼠中没有(图2e)。
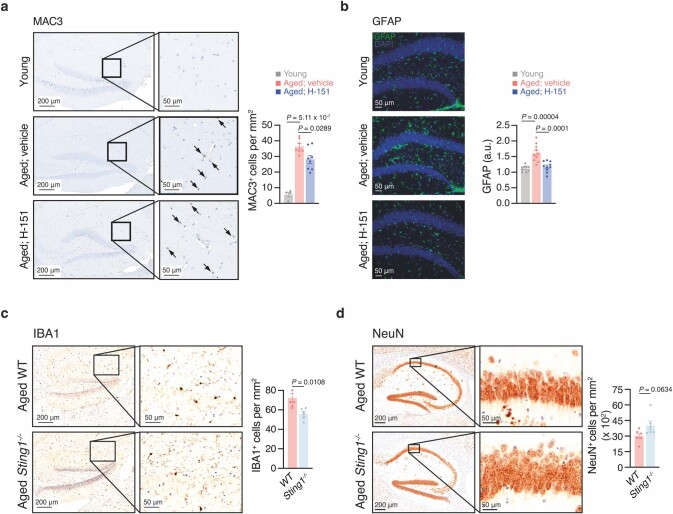
a,年轻小鼠(n = 4)和老龄小鼠(n = 7)或H-151处理小鼠(n = 8)脑切片海马MAC3的代表性图像和定量。箭头表示MAC3细胞。比例尺,200 μm(左),50 μm(右)。
b,给药或H-151处理的年轻小鼠(n = 9)和老年小鼠(n = 10)脑切片海马GFAP的代表性图像和定量。比例尺,50 μm;A.u,任意单位。
c,d,老龄WT CA1区海马IBA (c)和NeuN (d)的代表性图像及定量(IBA1, n = 5;NeuN, n = 6)和Sting1+ - / -小鼠(n = 6)。比例尺,200 μm(左),50 μm(右)。
数据为平均值±s.e.m。P值通过单因素方差分析计算,然后进行Tukey多重比较检验(a,b)或双侧Student’s unpaired t检验(c,d)。同时,当STING被抑制时,衰老的小胶质细胞溶酶体标志物MAC3的表达减少(扩展图5a)。H-151也能降低老年小鼠海马星形胶质细胞的免疫反应性(扩展图5b)。
我们证实,与衰老的野生型小鼠相比,衰老的Sting1−/−小鼠海马区的小胶质细胞积累减少,神经元密度增加(扩展图5c,d)。因此,这些结果表明,STING影响老年小鼠的大脑稳态。
综上所述,这些数据表明,在小鼠衰老的大脑中,STING被激活,并暗示在衰老过程中,STING信号上游的cGAS活性异常。
老年小胶质细胞的STING活性
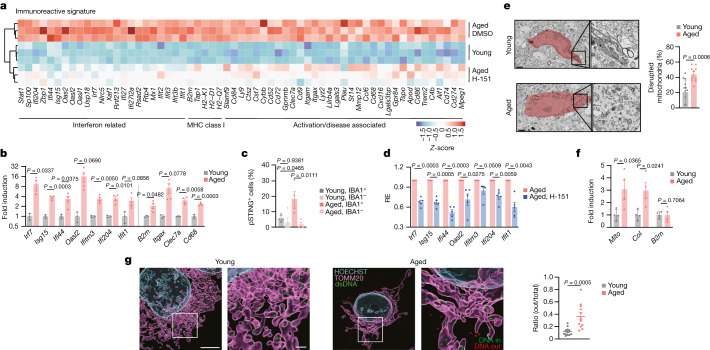
a,使用或不使用H-151处理的幼龄和老年小鼠海马体RNA-seq分析的差异基因表达(n = 3)。热图显示了显著DEG总列表中的I型IFN、MHC I类和小胶质细胞激活/疾病相关基因(扩展图6a;假发现率(FDR)≤0.01,|log2[fold change (FC)]|≥0.6)。
b,从幼龄(n = 3)和老年(n = 4)小鼠分离的初级小胶质细胞中免疫反应基因、ISGs和激活标记物的mRNA表达水平。
c,青年和老年海马切片pSTING染色的共聚焦成像定量(平均每只小鼠100-200个细胞,n = 4), IBA1 -和IBA1细胞差异定量。
d,老龄小鼠小胶质细胞中免疫反应性基因和ISGs mRNA表达水平。n = 5。
e,年轻(n = 13)和老年(n = 10)细胞中与年龄相关的小胶质细胞形态差异和每个海马小胶质细胞中线粒体破坏百分比的透射电镜图像,每种情况下随机选择3只小鼠。比例尺,1 μm。
f,年轻和年老小鼠小胶质细胞中Mito(线粒体DNA序列)、CoI和B2m的胞浆表达水平(n = 4)。
g,从年轻和老年小鼠分离的小胶质细胞线粒体外细胞质DNA病灶的代表性三维重建和定量。测量每个细胞线粒体外的DNA病灶相对于细胞质病灶(内部(绿色)和外部(红色))总数的比例。N = 12个细胞,来自3只小鼠。比例尺,上为5 μm,下为1 μm。
数据为平均值±s.e.m。P值的计算采用双侧未配对学生t检验(b和d-g)和单向方差分析,然后采用Tukey多重比较检验(c)。Mito指的是线粒体(mitochondria);CoI代表线粒体呼吸链复合物I(Complex I);B2m代表β2微球蛋白(beta-2 microglobulin)。
为了揭示大脑中cGAS-STING激活的潜在机制,我们对年轻成年小鼠和未接受H-151治疗的老年小鼠的海马进行了大量RNA测序(RNA-seq)分析。转录分析显示,在年轻海马和老年海马之间存在459个显著差异表达的基因(DEGs),其中很大一部分基因与先天免疫有关,包括与I型IFN信号传导和小胶质细胞功能相关的基因(图3a,扩展图6a)。H-151组对几种DEG的诱导减弱,并与幼鼠聚集在一起(图3a,扩展图6a)。从老年小鼠大脑中分离的初级小胶质细胞在形成CNS的免疫状态方面发挥着核心作用,其先天免疫基因的表达也出现了类似的增加(图3b)。因此,我们认为STING在脑老化中的神经退行性作用涉及小胶质细胞。为了研究小胶质细胞内的STING反应,我们可视化了磷酸化的STING (Ser365;pSTING)是海马区STING活动的直接标志。与年轻小鼠相比,pSTING病灶在老年海马中富集,并且大部分pSTING信号来自IBA1小胶质细胞(图3c, 扩展图6b,c)。老龄小胶质细胞的离体实验证实了STING依赖性I型IFN和促炎基因的表达(图3d)。
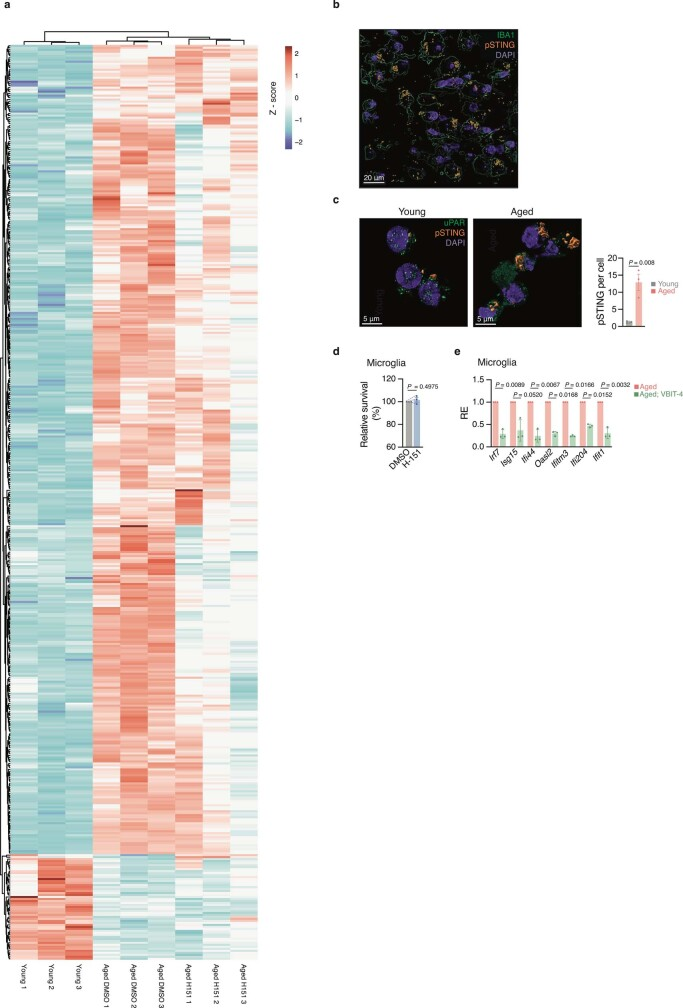
a,衰老反应基因热图(459个基因,FDR≤0.01,Log2FC≥0.6 n = 3)。颜色代表每百万的标准化计数。基因和样本通过无监督聚类排序。
b,老龄海马pSTING染色的代表性共聚焦图(n = 1,代表n = 4只小鼠)。标尺,20 μm。
c,青年和老年小鼠培养的小胶质细胞中具有代表性的共聚焦图像和pSTING灶的定量(平均每只小鼠20个细胞,n = 3)。比例尺,5 μm。
d,用细胞滴度蓝(CTB)法测定h -151处理24 h的初代小胶质细胞的相对存活率(n = 3只小鼠)。
e, VBIT-4 (10 μm)处理或未处理4天老龄小鼠原代小胶质细胞中免疫反应性和干扰素刺激基因的mRNA表达水平(n = 3)。
数据为平均值±s.e.m。P值采用双侧Student’s unpaired (c,d)或双侧Student’s paired (e) t检验计算。尿激酶型纤溶酶原激活物受体。VBIT-4是一种蛋白质,它是DNA感知的一个重要组成部分。
总的来说,这些结果证明了老年小鼠小胶质细胞中的STING活性,它指导了老化大脑中的先天免疫激活。
mtDNA激活老年小胶质细胞中的cGAS
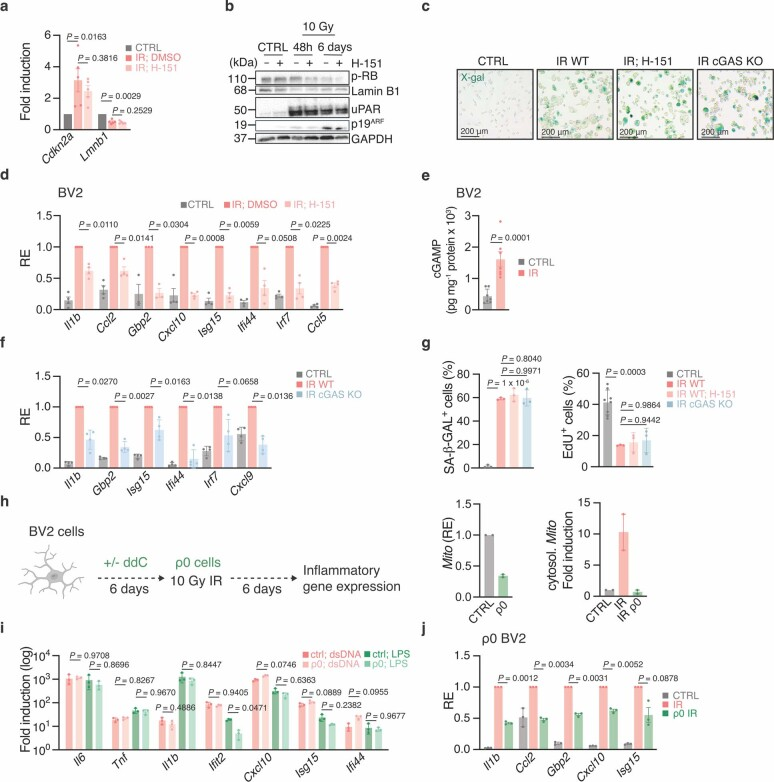
a-d, BV2细胞辐照(10 Gy, IR),然后DMSO-或H-151处理(每天,1 μM,照射后4-6天)。衰老标志物mRNA表达水平(n = 5个实验)(a)、Western blot表征(b)和衰老相关-β-半乳糖苷酶染色(n = 1,代表n = 3个实验)(c)。比尺,200 μm。每次实验(n = 4)测量对照和辐照DMSO或H-151处理细胞中促炎和干扰素刺激基因的mRNA表达水平(d),相对于辐照DMSO处理细胞。
e,对照和辐照BV2细胞裂解物中的cGAMP水平(n = 7个实验)。
f,对照和辐照的WT和cGAS-KO BV2细胞中促炎基因和干扰素刺激基因的mRNA表达水平(n = 4个实验)。
g,衰老相关-β-半乳糖苷酶(左,n = 3 FOV)和% EdU细胞(右)来自对照(n = 8 FOV)和辐照(n = 3 FOV) WT BV2, H-151处理或未处理如(a)和cGAS-KO BV2 (n = 3 FOV),代表n = 3个实验。
h,分析mtDNA缺失的BV2细胞(ρ0 BV2)的实验装置(左)。ρ0 BV2全细胞裂解液(中)和辐照后细胞质中(右)线粒体DNA序列Mito水平(n = 2次实验)。
i、LPS或dsDNA 90mer刺激的ρ0 BV2细胞促炎基因和干扰素刺激基因mRNA表达水平(n = 3个实验)。
j,对照和辐照的ρ0 BV2细胞中促炎和干扰素刺激基因的mRNA表达水平,每次实验(n = 3)相对于辐照未处理(IR)细胞。
数据为平均值±s.e.m。P值采用双侧配对比例Student’s t检验(a,d,e,j)、双侧未配对Student’s t检验(f)和单因素方差分析后的Tukey’s多重比较检验(g,i)获得。uPAR:尿激酶型纤溶酶原激活物受体。线粒体DNA (mtDNA)是cGAS-STING信号传导的中心激活因子,线粒体稳态破坏是衰老和神经退行性疾病的标志。透射电镜下,老龄小胶质细胞线粒体出现畸形,缺乏其特有的内部结构(图3e)。为了确定mtDNA是否释放到衰老小胶质细胞的细胞质中,我们采用反转录定量PCR (RT-qPCR)分析了原代小胶质细胞的细胞质DNA含量。与年轻小鼠的小胶质细胞相比,衰老的小胶质细胞显示mtDNA丰度增加,但基因组DNA种类没有增加(图3f)。老年小胶质细胞的超分辨率Airyscan成像证实,线粒体外膜附近有明显的mtDNA类核积聚,而在年轻小胶质细胞的细胞质中基本没有(图3h)。接下来,我们试图解决异常mtDNA是否有助于衰老小胶质细胞的炎症表型。先前的研究表明,电压依赖性离子通道(VDAC) 1/3低聚物参与mtDNA的胞质释放。VDAC寡聚化抑制剂VBIT-4体外治疗衰老小胶质细胞,抑制了几种I型IFN和促炎基因(扩展图6e)。体外细胞衰老模型证实了mtDNA在小胶质细胞来源的小鼠BV2细胞中触发cGAS依赖性炎症反应的作用(扩展图7)。
总之,这些数据表明mtDNA在指导衰老小胶质细胞炎症反应中的作用,并定义了衰老和衰老过程中cGAS-STING激活的分子模型。
小胶质细胞cGAS对大脑的影响
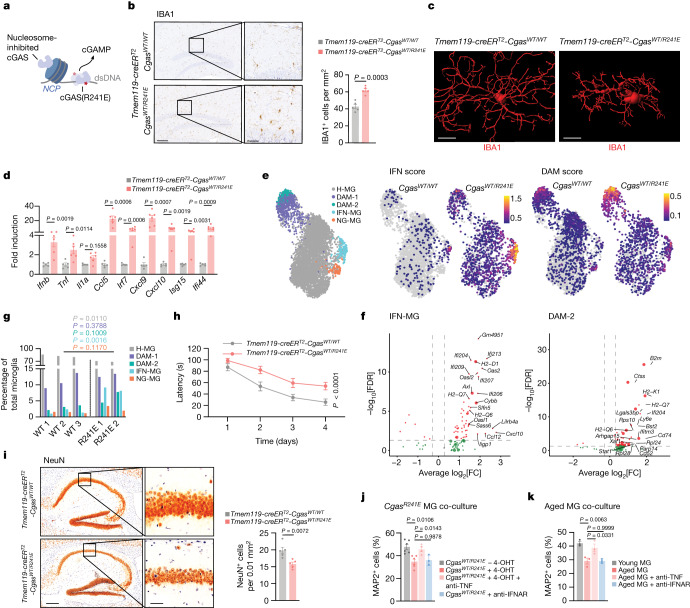
a,核小体结合缺陷cGAS突变体激活示意图。
b, Tmem119-creER^{T2}-Cgas^{WT/WT}和Tmem119-creER^{T2}-Cgas^{WT/R241E}小鼠海马IBA1染色的代表性图像和定量。n = 5。比例尺,左为200 μm,右为50 μm。
c, IMARIS重建的典型IBA1小胶质细胞。标尺,10 μm。
d, Tmem119-creER^{+T2}-Cgas^{WT/WT} (n = 5)和Tmem119-creER^{T2}-Cgas^{WT/R241E} (n = 6)小鼠脑促炎基因和ISGs mRNA表达水平。
e,均匀流形近似和投影(UMAP)图显示小胶质细胞单个核(snRNA-seq),以细胞身份着色(左,稳态小胶质细胞(H-MG);疾病相关小胶质细胞(DAM-1/2);IFN相关小胶质细胞(IFN-MG);神经退行性相关小胶质细胞(ND-MG))和IFN/DAM基因表达评分按Cgas基因型划分(右)。彩色刻度条表示基因负荷评分。
f, IFN-MG和DAM-2-MG中Cgas^{WT/WT}和Cgas^{WT/R241E}的差异(FDR≤0.05,log2[FC]≥0.3)。超大点表示与相关状态相关的基因(补充表6)。
g,通过snRNA-seq分析Tmem119-creER^{T2}-Cgas^{WT/WT} (WT, n = 3)和Tmem119-creER^{T2}-Cgas^{WT/R241E} (R241E, n = 2)小胶质细胞群体的相对比例。
h, Tmem119-creER^{T2}-Cgas^{WT/WT} (n = 6)和Tmem119-creER^{T2}-Cgas^{WT/R241E} (n = 11)小鼠Morris水迷宫实验。p = 1 × 10−5。
i, Tmem119-creER^{+T2}-Cgas^{WT/WT}和Tmem119-creER^{T2}-Cgas^{WT/R241E}小鼠海马CA1区NeuN细胞的代表性图像和定量。n = 5。比例尺,左250 μm,右50 μm。
j,k,用Rosa26-creER^{+T2}-Cgas^{WT/R241E}分离的小胶质细胞培养MAP2神经元的相对存活率,分别用4-OHT (n = 6)和TNF-中和抗体(n = 4)或IFNAR-中和抗体(n = 3载片)处理n = 3只小鼠(j);或用TNF-或IFNAR中和抗体处理的年轻和年老小鼠的小胶质细胞(k)(平均每只小鼠,n = 3)。
数据为平均值±s.e.m。P值的计算采用双侧Student’s unpaired t检验(b, d, g和i),单因素方差分析,然后采用Tukey’s多重比较检验(j和k)和普通双因素方差分析(h)。Tmem119-creER 是一种基因表达载体,可以在特定细胞类型中启动特定基因的表达。Cgas 表示该转基因小鼠携带cGAS基因的一个突变,即在第241位氨基酸上发生了替换,将精氨酸(R)替换为谷氨酸(E)。IMARIS是一种三维图像分析软件,常用于细胞和组织的图像处理和重建。snRNA-seq (single-nucleus RNA sequencing) 是一种针对细胞核中的RNA进行测序的技术。它主要应用于那些难以获得完整细胞或细胞团的样本,如神经组织和冷冻保存的组织。
不同的免疫信号机制涉及神经退行性疾病,可能与cGAS-STING信号轴协同促进年龄相关的神经退行性疾病。为了确定单独参与cGAS-STING通路是否足以驱动神经病理特征,我们设计了一种遗传策略来触发体内cGAS活性。之前,我们和其他人发现,cGAS通过与核小体结合而受到抑制,核小体调节的破坏使cGAS具有强大的DNA依赖性激活(图4a)。
此外,他莫昔芬处理的mg-Cgas小鼠大脑中炎症基因和I型IFN相关基因水平升高(图4d)。
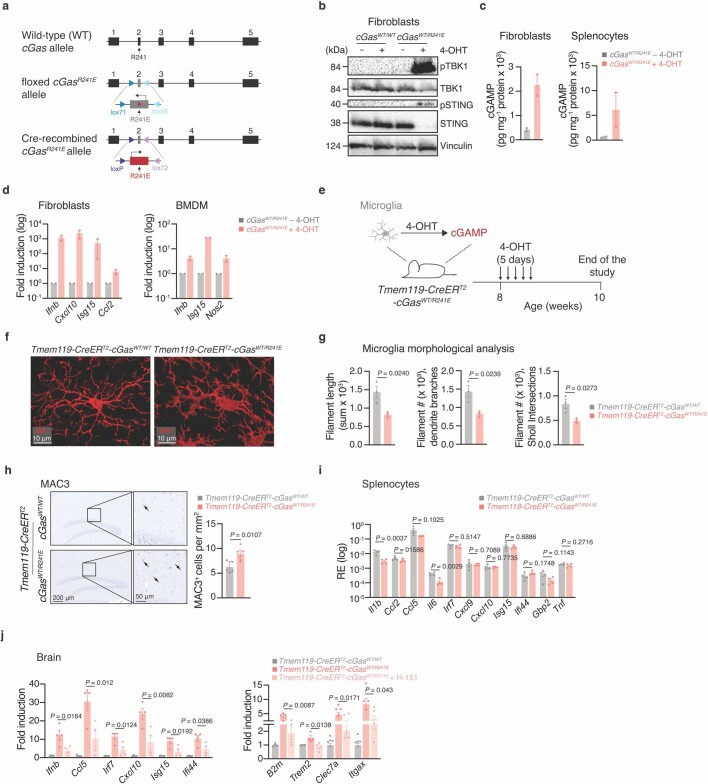
a, 敲入cGAS功能获得小鼠模型示意图。
b, Western blot分析4-OHT处理或未处理的Rosa26-creER^{T2}-cGas^{WT/WT}和Rosa26-creER^{T2}-cGas^{WT/R241E}小鼠尾端成纤维细胞中cGAS的活化情况。
c, Rosa26-creER^{T2}-cGas^{WT/WT}和Rosa26-creER^{T2}-cGas^{WT/R241E}小鼠尾端成纤维细胞和脾细胞裂解液中的cGAMP水平(n = 2)。
d,干扰素刺激基因在Rosa26-creER^{T2}-cGas^{WT/R241E}小鼠(n = 2)尾尖成纤维细胞和骨髓源性巨噬细胞(BMDM)中的mRNA表达水平,4-OHT处理或未处理。
e,用于分析小胶质细胞特异性cGAS激活的实验装置。
f, g,通过IMARIS分析Tmem119-creER^{+T2}-cGas^{WT/WT}和Tmem119-creER^{T2}-cGas^{WT/R241E}小鼠的IBA1小胶质细胞,显示原始图像预重建(f)和形态学分析(g),并基于图像量化(n = 3只小鼠)。
h, Tmem119-creER^{T2}-cGas^{WT/WT}或Tmem119-creER^{T2}-cGas^{WT/R241E}小鼠脑切片海马MAC3的代表性图像及定量(n = 5)。
i, 4-OHT处理Tmem119-creER^{T2}-cGas^{WT/WT}和Tmem119-creER^{T2}-cGas^{WT/R241E}小鼠脾细胞中促炎基因和干扰素刺激基因的mRNA表达水平(n = 4)。
j, Tmem119-creER^{T2}-cGas^{WT/WT}和Tmem119-creER^{T2}-cGas^{WT/R241E}小鼠脑促炎和干扰素刺激基因的mRNA表达水平(左)和小胶质细胞激活标志物(右),4-OHT处理小鼠,以及不加H-151处理小鼠(n = 4)。
数据为平均值±s.e.m。P值采用双侧Student’s unpaired t检验(g-i)或单因素方差分析,然后采用Tukey’s多重比较检验(j)。RE为相对表达。我们生成的小鼠中,核小体结合突变体Cgas(对应于人类CGAS)可以通过Cre重组酶有条件地表达(扩展图8a)。在体外,4-羟基他莫昔芬(4-OHT)处理具有他莫昔芬诱导的Rosa26-creER等位基因的小鼠细胞,导致cGAMP合成和I型IFN应答基因上调,证明Cgas小鼠可以用于研究体内cGAS激活的影响(扩展图8b-d)。
我们生成了Tmem119-creER-Cgas小鼠(以下简称小胶质(mg)-Cgas),允许他莫昔芬诱导的Cgas主要局限于小胶质细胞和脑巨噬细胞的表达(扩展图8e)。在给予他莫昔芬2周后,与对照组小鼠相比,mg-Cgas小鼠在包括海马在内的几个脑区显示出具有激活形态的小胶质细胞数量的强烈增加(图4b,c和扩展图8f-h)。在脾脏等外周器官中未检测到炎症标记基因的升高,排除了脑外部过程对观察到的变化的贡献(扩展图8i)。正如预期的那样,H-151处理减弱了mg-Cgas小鼠的小胶质细胞激活(扩展图8j)。

a,富集小胶质细胞的无监督聚类的UMAP图(25个单独的簇),小胶质细胞特异性表达评分,将簇0-4和17识别为小胶质细胞,以及过滤小胶质细胞的无监督聚类。
b,个体小鼠间与小胶质细胞状态相关的基因表达评分小提琴图。
c,在DAM和IFN亚群中,cGasWT/R241E和cGasWT/WT小鼠之间相关基因和差异表达的代表性UMAP图。
d, cGas^{WT/R241E}和cGas^{WT/WT}小胶质细胞与来自(Tabula Muris, GSM4505405)的24m龄小胶质细胞的整合。IFN-MG和Louvain cluster 4 (IFN相关的衰老小胶质细胞)、DAM-1/2-MG和ND-MG的UMAP图、小鼠年龄和小胶质细胞相关亚群基因的基因表达评分显示,IFN群体和DAM/ND群体与表达高水平DAM和神经退行性(ND)标志物的衰老小胶质细胞共聚类。
e, cGas^{WT/R241E}和cGas^{WT/WT}小胶质细胞与疾病相关小胶质细胞的整合(Sala Frigerio, GSE127892, F_IFN:干扰素应答小胶质细胞,F_ARM:激活反应小胶质细胞,F_TRM:过渡反应小胶质细胞,F_MHC-ARM:主要组织相容性高ARMs, F_MG:稳态小胶质细胞)。IFN-MG和F_IFN、DAM-1/2-MG和F_ARM、F_TRM、F_MHC-ARM、MG和F_MG/2的UMAP图以及小胶质细胞相关亚群基因表达评分显示,IFN群体和DAM/TRM群体与表达高水平IFN和DAM标记的疾病相关小胶质细胞共聚。为了全面定义cGAS控制的小胶质细胞转录程序,我们对9505个富集小胶质细胞进行了单核RNA-seq (snRNA-seq)分析,从中我们从他莫昔芬处理的mg-Cgas小鼠和对照Cgas小鼠中获得的5360个小胶质细胞进行了生物信息学鉴定(扩展图9a)。聚类分析揭示了三种不同的小胶质细胞转录状态,显示出一种独特的转录谱,将它们与主要的稳态小胶质细胞群体区分开来(图4e和图9b,c)。这些转录簇对应于先前确定的亚群,包括疾病相关(DAM)、IFN相关和与衰老和神经退行性疾病相关的神经退行性小胶质状态(扩展数据图9d,e)。来自这三种状态的细胞都强烈上调与IFN信号(Ifits, Oasl1, Oasl2, Isg15, Stat2), DAM信号(B2m, Ctsb, Ctsd, Ctsz, Cd9, Cd63)和神经退行性信号(Apbb2, Olfr111, Xylt1)相关的基因(补充表3)。此外,在DAM亚型中,我们观察到从低激活状态到高激活状态(DAM-1到DAM-2)的进展。mg-Cgas细胞处于高激活状态的比例更高(图4e、g和扩展图9b)。两种小鼠模型之间的比较显示,mg-Cgas细胞中与IFN和DAM状态相关的基因(最明显的是在DAM-2群体中)的扩增和更高的表达,以及IFN信号在全球小胶质群体中的普遍增加(图4e,g、扩展图9b,c和补充表4)。
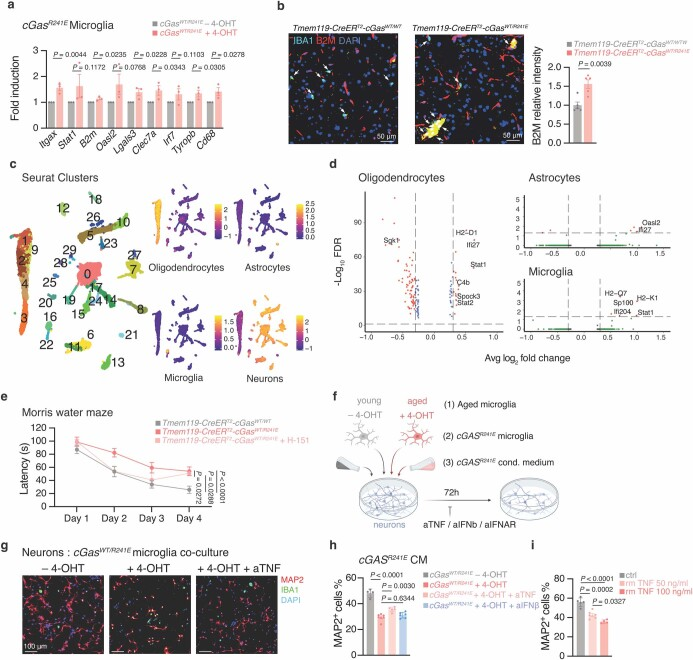
a,干扰素刺激基因和激活标记在Tmem119-creER^{T2}-cGas^{WT/R241E}小鼠小胶质细胞中的mRNA表达水平(n = 3)。
b, Tmem119-creER^{+T2}-cGas^{WT/WT}和Tmem119-creER^{T2}-cGas^{WT/R241E}小鼠脑切片IBA1小胶质细胞B2M强度的代表性图像和定量(平均每只小鼠,n = 5, 10-15个细胞)。箭头表示IBA小胶质细胞。标尺,50 μm。
c,分类海马核的无监督聚类(38簇)的UMAP图,具有鉴定的少突胶质细胞,星形胶质细胞,小胶质细胞和神经元特异性基因表达评分。
d,火山地块之间的基因差异表达Tmem119-creER^{+T2}-cGAS^{WT/WT}和Tmem119-creER^{T2}-cGAS^{WT/R241E}老鼠(n = 4)少突胶质细胞、星形胶质细胞、小胶质细胞和MG(RDF≤0.05,Log2FC≥0.3,基因补充表6中列出)。
e,莫里斯水迷宫的记忆测试Tmem119-creER^{T2}-cGAS^{WT/WT}(n = 6)和Tmem119-creER^{T2}-cGAS^{WT/R241E}老鼠在图4h),此外含H-151(n = 5) 不含(n = 11), P = 1×10^−5。
f, 神经细胞共培养实验示意图。
g, Rosa26-creER^{T2}-cGas^{WT/R241E}小鼠(IBA1,绿色)小胶质细胞培养的神经元(MAP2,红色)、4-OHT处理或未处理的神经元和aTNF的代表性图像(代表n = 3个实验)。标尺,100 μm。
h, Rosa26-creER^{+T2}-cGas^{WT/R241E}分离巨噬细胞条件培养基培养MAP2神经元的相对存活率,4-OHT处理(n = 6 FOV)或未处理(n = 4 FOV),以及添加aTNF (n = 5 FOV), aIFNb (n = 6 FOV,来自n = 3只小鼠),P = 3 × 10^−9。
i,增加重组TNF剂量(未处理n = 5, 50 ng/ml n = 5, 100 ng/ml n = 4 FOV)处理MAP2神经元的相对存活率,每种情况3张载玻片,P = 1 × 10^−5。
数据为平均值±s.e.m。P值通过单侧(a)或双侧(b)未配对学生t检验、单因素方差分析、多因素比较检验(h,i)或双因素方差分析(e)获得。我们在表达Cgas的分离小胶质细胞中证实了I型IFN-和DAM相关基因的上调,并证实了mg-Cgas小鼠大脑中B2M小胶质细胞的比例增加(扩展图10a,b)。
因此,这些数据表明,在缺乏额外触发因素的情况下,仅cGAS就足以促进小胶质细胞向独特的衰老和疾病相关激活状态过渡。
为了解决cGAS在小胶质细胞中的激活如何影响大脑中的其他细胞群,我们对mg-Cgas小鼠的海马进行了snRNA-seq分析,这些小鼠接受或未接受他莫昔芬治疗。对每组4只小鼠收集的21,500个细胞进行聚类分析,揭示了四种主要的脑细胞类型,包括小胶质细胞、星形胶质细胞、少突胶质细胞和神经元(扩展图10c,d)。在少突胶质细胞和星形胶质细胞中,小胶质细胞cGAS的激活伴随着IFN相关的转录组变化,而神经元则表现出最小的转录变化(扩展数据图10c、d和补充表5)。在少突胶质细胞中,差异表达最多的基因是在衰老和神经退行性变过程中表征这种细胞类型的几个基因,包括H2-D1、if27、C4b、Spock3、Trf和Sgk1。
因此,除了小胶质细胞的细胞内在作用外,cGAS激活还指导非免疫胶质细胞的转录程序,类似于老年大脑中发生的转录程序。
cGAS激活驱动神经变性
为了确定小胶质细胞中的cGAS活性是否可以启动神经退行性过程,我们将海马依赖性学习和海马神经元密度作为大脑可塑性的两个独立指标进行了研究。与对照组相比,mg-Cgas小鼠在Morris水迷宫测试中表现出认知能力受损,同时海马神经元密度下降(图4h,i)。与衰老过程一样,H-151减轻了mg-Cgas小鼠学习能力的退化,这与STING的促炎作用影响神经元功能的模型一致(扩展图10e)。为了解剖小胶质细胞cGAS-STING活性下游的神经毒性过程,我们用原代野生型神经元和Cgas小鼠的小胶质细胞进行了共培养实验(扩展数据图10f)。cGAS活性有效地抑制了神经元细胞的存活,这种作用也在cgas表达细胞的条件培养基中得到了再现,表明cGAS介导的神经毒性涉及可溶性介质的分泌(图4j和扩展图10g,h)。我们的snRNA小胶质转录组数据分析显示,相对于稳态小胶质细胞,Tnf在IFN相关的小胶质细胞状态中上调,并在mg-Cgas小鼠的大脑中被诱导(图4d和补充表3)。值得注意的是,添加一种针对TNF的中和抗体,可以强有力地挽救Cgas诱导的神经元死亡(图4j和扩展图10h)。相比之下,阻断I型IFN信号没有效果(图4j和扩展图10h)。与衰老小胶质细胞的共培养实验证实了TNF在连接异常cGAS活性与受损神经元细胞存活之间的关键作用(图4k和扩展图10i)。
3讨论
在这里,我们建立了cGAS-STING通路作为衰老相关炎症的重要驱动因素,并确定了cGAS-STING在衰老相关神经变性建立中的小胶质内参与。在脑细胞中,特别是在小胶质细胞中,I型IFN标记的存在越来越多地与各种物种的衰老和神经退行性疾病相关。我们对自然衰老小鼠的研究表明,cGAS-STING信号的激活是小胶质细胞中与衰老相关的I型IFN反应的重要因素,可直接导致神经元丢失和认知障碍。cGAS功能获得小鼠的单细胞转录谱进一步揭示,除了I型IFN外,cGAS单独激活会触发许多神经退行性疾病状态和衰老过程中共享的核心小胶质基因表达程序。因此,除了自然衰老之外,这些结果指出小胶质细胞cGAS-STING激活在退行性脑疾病中具有更广泛的作用。
体外共培养研究表明,TNF是cGAS控制的小胶质细胞反应的关键神经毒性因子。此外,我们的snRNA-seq分析发现,少突胶质细胞和星形胶质细胞的激活状态发生了相当大的继发性变化,它们可以作为传播神经毒性信号的重要中间细胞。阐明影响cGAS下游小胶质细胞依赖性神经毒性的精确神经免疫串扰,将有助于深入了解它们在神经退行性疾病中的作用,是未来研究的一个重要领域。
我们对活化小胶质细胞的机制表征表明,线粒体dna在细胞质中积累以刺激衰老细胞,从而在衰老的两个主要特征(即线粒体功能障碍和炎症)之间提供功能联系。先前的研究表明,DNA损伤产物和逆转录因子扩增是衰老过程中cGAS的上游触发因素。我们假设,在衰老过程中,就像在其他复杂疾病中一样,不同来源的DNA可以协同作用,促进cGAS活性。我们提出异常mtDNA也有助于外周衰老相关的炎症现象,这是由cGAS-STING级联控制的。
伴随神经退行性变的炎症反应的损伤相关触发器可能以疾病特异性的方式产生。结合先前对阿尔茨海默病、帕金森氏病、肌萎缩侧索硬化症和额颞叶痴呆以及尼曼-皮克病模型的研究,我们的研究揭示了cGAS-STING信号在慢性神经退行性疾病中的显著趋同。仔细观察,免疫激活的分子(基因组与mtDNA)和细胞(小胶质细胞与神经元)起源方面出现差异,这可能会对疾病本身的特征产生影响。
原文:Gulen MF, Samson N, Keller A, et al. cGAS-STING drives ageing-related inflammation and neurodegeneration. Nature. 2023;620(7973):374-380. doi:10.1038/s41586-023-06373-1
声明:文中观点不代表本站立场。本文传送门:https://eyangzhen.com/360991.html
